Что такое биосинтез днк
Содержание:
- Транскрипция у эукариот
- Трансляция
- Суперскрученная молекула
- Транскрипция и трансляция у прокариот
- Строение и функции ДНК
- ДНК (дезоксирибонуклеиновая кислота)
- Окончательная компактизация ДНК
- Как происходит репликация
- Трансляция
- Сущность генетического кода
- Как воспроизводится генетическая программа
- Функции ДНК
- Транскрипция и трансляция
- Обмен веществ
- Нуклеиновые кислоты – полимерные молекулы
- Строение и функции РНК
- А-ДНК – сухая молекула
- Механизм транскрипции
- Основные аспекты синтеза молекул на основе матрицы
- Репликация ДНК и ее строение
Транскрипция у эукариот
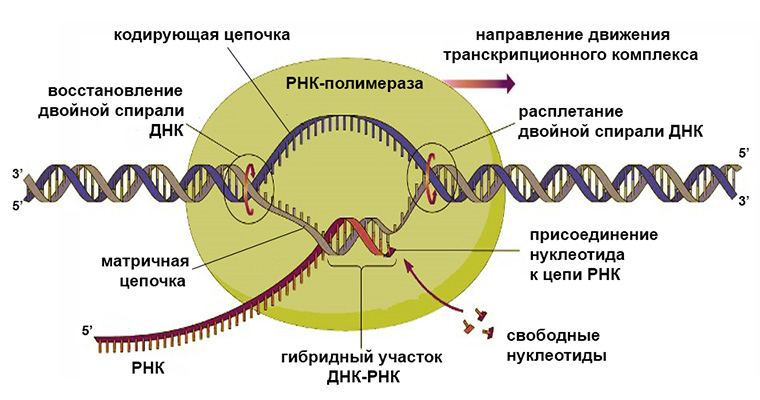
Транскрипция — синтез РНК на матрице ДНК. Осуществляется ферментом РНК-полимеразой.
РНК-полимераза может присоединиться только к промотору, который находится на 3′-конце матричной цепи ДНК, и двигаться только от 3′- к 5′-концу этой матричной цепи ДНК. Синтез РНК происходит на одной из двух цепочек ДНК в соответствии с принципами комплементарности и антипараллельности. Строительным материалом и источником энергии для транскрипции являются рибонуклеозидтрифосфаты (АТФ, УТФ, ГТФ, ЦТФ).
В результате транскрипции образуется «незрелая» иРНК (про-иРНК), которая проходит стадию созревания или процессинга. Процессинг включает в себя: 1) КЭПирование 5′-конца, 2) полиаденилирование 3′-конца (присоединение нескольких десятков адениловых нуклеотидов), 3) сплайсинг (вырезание интронов и сшивание экзонов). В зрелой иРНК выделяют КЭП, транслируемую область (сшитые в одно целое экзоны), нетранслируемые области (НТО) и полиадениловый «хвост».
Транслируемая область начинается кодоном-инициатором, заканчивается кодонами-терминаторами. НТО содержат информацию, определяющую поведение РНК в клетке: срок «жизни», активность, локализацию.
Транскрипция и процессинг происходят в клеточном ядре. Зрелая иРНК приобретает определенную пространственную конформацию, окружается белками и в таком виде через ядерные поры транспортируется к рибосомам; иРНК эукариот, как правило, моноцистронны (кодируют только одну полипептидную цепь).
Трансляция
Трансляция — синтез полипептидной цепи на матрице иРНК.
Органоиды, обеспечивающие трансляцию, — рибосомы. У эукариот рибосомы находятся в некоторых органоидах — митохондриях и пластидах (70S-рибосомы), в свободном виде в цитоплазме (80S-рибосомы) и на мембранах эндоплазматической сети (80S-рибосомы). Таким образом, синтез белковых молекул может происходить в цитоплазме, на шероховатой эндоплазматической сети, в митохондриях и пластидах. В цитоплазме синтезируются белки для собственных нужд клетки; белки, синтезируемые на ЭПС, транспортируются по ее каналам в комплекс Гольджи и выводятся из клетки. В рибосоме выделяют малую и большую субъединицы. Малая субъединица рибосомы отвечает за генетические, декодирующие функции; большая — за биохимические, ферментативные.
В малой субъединице рибосомы расположен функциональный центр (ФЦР) с двумя участками — пептидильным (Р-участок) и аминоацильным (А-участок). В ФЦР может находиться шесть нуклеотидов иРНК, три — в пептидильном и три — в аминоацильном участках.
Для транспорта аминокислот к рибосомам используются транспортные РНК, тРНК (лекция №4). Длина тРНК от 75 до 95 нуклеотидных остатков. Они имеют третичную структуру, по форме напоминающую лист клевера. В тРНК различают антикодоновую петлю и акцепторный участок. В антикодоновой петле РНК имеется антикодон, комплементарный кодовому триплету определенной аминокислоты, а акцепторный участок на 3′-конце способен с помощью фермента аминоацил-тРНК-синтетазы присоединять именно эту аминокислоту (с затратой АТФ). Таким образом, у каждой аминокислоты есть свои тРНК и свои ферменты, присоединяющие аминокислоту к тРНК.
|
1 — антикодон;2 — участок, связывающий аминокислоту. |
Транспорт аминокислот к рибосомам: 1 — фермент; 2 — тРНК; 3 — аминокислота. |
Двадцать видов аминокислот кодируются 61 кодоном, теоретически может быть 61 вид тРНК с соответствующими антикодонами. Но кодируемых аминокислот всего 20 видов, значит, у одной аминокислоты может быть несколько тРНК. Установлено существование нескольких тРНК, способных связываться с одним и тем же кодоном (последний нуклеотид в антикодоне тРНК не всегда важен), поэтому в клетке обнаружено всего около 40 различных тРНК.
Синтез белка начинается с того момента, когда к 5′-концу иРНК присоединяется малая субъединица рибосомы, в Р-участок которой заходит метиониновая тРНК (транспортирующая аминокислоту метионин). Следует отметить, что любая полипептидная цепь на N-конце сначала имеет метионин, который в дальнейшем чаще всего отщепляется. Синтез полипептида идет от N-конца к С-концу, то есть пептидная связь образуется между карбоксильной группой первой и аминогруппой второй аминокислот.
Затем происходит присоединение большой субъединицы рибосомы, и в А-участок поступает вторая тРНК, чей антикодон комплементарно спаривается с кодоном иРНК, находящимся в А-участке.
Пептидилтрансферазный центр большой субъединицы катализирует образование пептидной связи между метионином и второй аминокислотой. Отдельного фермента, катализирующего образование пептидных связей, не существует. Энергия для образования пептидной связи поставляется за счет гидролиза ГТФ.
Как только образовалась пептидная связь, метиониновая тРНК отсоединяется от метионина, а рибосома передвигается на следующий кодовый триплет иРНК, который оказывается в А-участке рибосомы, а метиониновая тРНК выталкивается в цитоплазму. На один цикл расходуется 2 молекулы ГТФ. В А-участок заходит третья тРНК, и образуется пептидная связь между второй и третьей аминокислотами.
Трансляция идет до тех пор, пока в А-участок не попадает кодон-терминатор (УАА, УАГ или УГА), с которым связывается особый белковый фактор освобождения. Полипептидная цепь отделяется от тРНК и покидает рибосому. Происходит диссоциация, разъединение субъединиц рибосомы.
Скорость передвижения рибосомы по иРНК — 5–6 триплетов в секунду, на синтез белковой молекулы, состоящей из сотен аминокислотных остатков, клетке требуется несколько минут. Первым белком, синтезированным искусственно, был инсулин, состоящий из 51 аминокислотного остатка. Потребовалось провести 5000 операций, в работе в течение трех лет принимали участие 10 человек.
В трансляции можно выделить три стадии: а) инициации (образование иницаторного комплекса), б) элонгации (непосредственно «конвейер», соединение аминокислот друг с другом), в) терминации (образование терминирующего комплекса).
Суперскрученная молекула
Сверхспирализованная форма ДНК возникает, когда двухцепочечная молекула совершает дополнительное закручивание. Оно может быть направлено по часовой стрелке (положительно) либо против (в этом случае говорят об отрицательной сверхспирализации). ДНК большинства организмов суперскручена отрицательно, то есть против основных витков «двойной спирали».
В результате образования дополнительных петель – супервитков – ДНК приобретает сложную пространственную конфигурацию. В клетках эукариот этот процесс происходит с формированием комплексов, в которых ДНК отрицательно навивается на гистоновые белковые комплексы и принимает вид нити с бусинами-нуклеосомами. Свободные участки нити называются линкерами. В поддержании суперскрученной формы молекулы ДНК принимают участие и негистоновые белки, а также неорганические соединения. Так образуется хроматин – вещество хромосом.
Хроматиновые нити с нуклеосомными бусинами способны к дальнейшему усложнению морфологии в процессе, называемом конденсацией хроматина.
Транскрипция и трансляция у прокариот
«Механизмы» сборки полинуклеотидных и полипептидных цепочек у прокариот и эукариот не различаются. Но в связи с тем, что гены прокариот не имеют экзонов и интронов (исключение — гены архебактерий), располагаются группами, и на эту группу генов приходится один промотор, появляются следующие особенности транскрипции и трансляции у прокариот.
- В результате транскрипции образуется полицистронная иРНК, кодирующая несколько белков, совместно обеспечивающих определенную группу реакций.
- иРНК имеет несколько центров инициации трансляции, терминации трансляции и НТО.
- Не происходят КЭПирование, полиаденилирование и сплайсинг иРНК.
- Трансляция начинается еще до завершения транскрипции; эти процессы не разделены во времени и пространстве, как это имеет место у эукариот.
1 — ДНК; 2 — РНК-полимераза; 3 — Нуклеозидтрифосфаты ГТФ, ЦТФ, АТФ, УТФ.
Можно добавить, что срок «жизни» прокариотических иРНК — несколько минут (у эукариот — часы и даже сутки).
-
Перейти к лекции №9 « Строение прокариотической клетки. Вирусы»
-
Перейти к лекции №11 « Понятие об обмене веществ. Биосинтез белков»
-
Смотреть оглавление (лекции №1-25)
Строение и функции ДНК
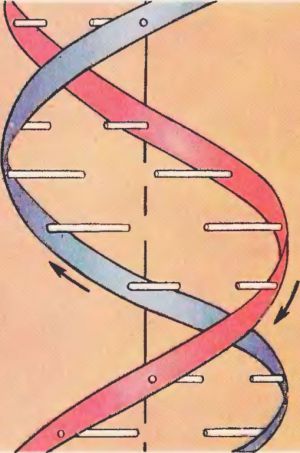
ДНК — полимер, мономерами которой являются дезоксирибонуклеотиды. Модель пространственного строения молекулы ДНК в виде двойной спирали была предложена в 1953 г. Дж. Уотсоном и Ф. Криком (для построения этой модели они использовали работы М. Уилкинса, Р. Франклин, Э. Чаргаффа).
Молекула ДНК образована двумя полинуклеотидными цепями, спирально закрученными друг около друга и вместе вокруг воображаемой оси, т.е. представляет собой двойную спираль (исключение — некоторые ДНК-содержащие вирусы имеют одноцепочечную ДНК). Диаметр двойной спирали ДНК — 2 нм, расстояние между соседними нуклеотидами — 0,34 нм, на один оборот спирали приходится 10 пар нуклеотидов. Длина молекулы может достигать нескольких сантиметров. Молекулярный вес — десятки и сотни миллионов. Суммарная длина ДНК ядра клетки человека — около 2 м. В эукариотических клетках ДНК образует комплексы с белками и имеет специфическую пространственную конформацию.
Мономер ДНК — нуклеотид (дезоксирибонуклеотид) — состоит из остатков трех веществ: 1) азотистого основания, 2) пятиуглеродного моносахарида (пентозы) и 3) фосфорной кислоты. Азотистые основания нуклеиновых кислот относятся к классам пиримидинов и пуринов. Пиримидиновые основания ДНК (имеют в составе своей молекулы одно кольцо) — тимин, цитозин. Пуриновые основания (имеют два кольца) — аденин и гуанин.
Моносахарид нуклеотида ДНК представлен дезоксирибозой.
Название нуклеотида является производным от названия соответствующего основания. Нуклеотиды и азотистые основания обозначаются заглавными буквами.
| Азотистое основание | Название нуклеотида | Обозначение |
|---|---|---|
| Аденин | Адениловый | А (A) |
| Гуанин | Гуаниловый | Г (G) |
| Тимин | Тимидиловый | Т (T) |
| Цитозин | Цитидиловый | Ц (C) |
Полинуклеотидная цепь образуется в результате реакций конденсации нуклеотидов. При этом между 3′-углеродом остатка дезоксирибозы одного нуклеотида и остатком фосфорной кислоты другого возникает фосфоэфирная связь (относится к категории прочных ковалентных связей). Один конец полинуклеотидной цепи заканчивается 5′-углеродом (его называют 5′-концом), другой — 3′-углеродом (3′-концом).
Против одной цепи нуклеотидов располагается вторая цепь. Расположение нуклеотидов в этих двух цепях не случайное, а строго определенное: против аденина одной цепи в другой цепи всегда располагается тимин, а против гуанина — всегда цитозин, между аденином и тимином возникают две водородные связи, между гуанином и цитозином — три водородные связи. Закономерность, согласно которой нуклеотиды разных цепей ДНК строго упорядоченно располагаются (аденин — тимин, гуанин — цитозин) и избирательно соединяются друг с другом, называется принципом комплементарности. Следует отметить, что Дж. Уотсон и Ф. Крик пришли к пониманию принципа комплементарности после ознакомления с работами Э. Чаргаффа. Э. Чаргафф, изучив огромное количество образцов тканей и органов различных организмов, установил, что в любом фрагменте ДНК содержание остатков гуанина всегда точно соответствует содержанию цитозина, а аденина — тимину («правило Чаргаффа»), но объяснить этот факт он не смог.
Из принципа комплементарности следует, что последовательность нуклеотидов одной цепи определяет последовательность нуклеотидов другой.
Цепи ДНК антипараллельны (разнонаправлены), т.е. нуклеотиды разных цепей располагаются в противоположных направлениях, и, следовательно, напротив 3′-конца одной цепи находится 5′-конец другой. Молекулу ДНК иногда сравнивают с винтовой лестницей. «Перила» этой лестницы — сахарофосфатный остов (чередующиеся остатки дезоксирибозы и фосфорной кислоты); «ступени» — комплементарные азотистые основания.
Функция ДНК — хранение и передача наследственной информации.
ДНК (дезоксирибонуклеиновая кислота)
ДНК (дезоксирибонуклеиновая кислота) – своеобразный чертеж жизни, сложный код, в котором заключены данные о наследственной информации. Эта сложная макромолекула способна хранить и передавать наследственную генетическую информацию из поколения в поколение.
ДНК определяет такие свойства любого живого организма как наследственность и изменчивость. Закодированная в ней информация задает всю программу развития любого живого организма. Генетически заложенные факторы предопределяют весь ход жизни человека.
Дезоксирибонуклеиновая кислота (ДНК) — макромолекула (одна из трёх основных, две другие — РНК и белки), обеспечивающая хранение, передачу из поколения в поколение и реализацию генетической программы развития и функционирования живых организмов. ДНК содержит информацию о структуре различных видов РНК и белков.
В клетках эукариот (животных, растений и грибов) ДНК находится в ядре клетки в составе хромосом, а также в некоторых клеточных органоидах (митохондриях и пластидах). В клетках прокариотических организмов (бактерий и архей) кольцевая или линейная молекула ДНК, так называемый нуклеоид, прикреплена изнутри к клеточной мембране. У них и у низших эукариот (например, дрожжей) встречаются также небольшие автономные, преимущественно кольцевые молекулы ДНК, называемые плазмидами.
ДНК — это длинная полимерная молекула, состоящая из повторяющихся блоков — нуклеотидов. Каждый нуклеотид состоит из азотистого основания, сахара (дезоксирибозы) и фосфатной группы.
Связи между нуклеотидами в цепи образуются за счёт дезоксирибозы и фосфатной группы (фосфодиэфирные связи).
В подавляющем большинстве случаев (кроме некоторых вирусов, содержащих одноцепочечную ДНК) макромолекула ДНК состоит из двух цепей.
Вторичная структура ДНК представляет собой двойную спираль, состоящую из двух параллельных неразветвленных полинуклеотидных цепей, закрученных в противоположные стороны вокруг общей оси.
Пуриновые и пиримидиновые основания расположены внутри спирали, а остатки фосфата и дезоксирибозы – снаружи.
Две спирали удерживаются вместе водородными связями между парами азотистых оснований. Водородные связи образуются между определенными основаниями: тимин (Т) образует водородные связи только с аденином (А), а цитозин (Ц) – только с гуанином (Г). В первой паре азотистых оснований две водородные связи, а во второй – три.
Такие пары оснований называются комплементарными парами. А такое пространственное соответствие молекул, способствующее их сближению и образованию водородных связей, называется комплементарностью. Комплементарность обусловливает спиралевидную модель ДНК.
Две спирали в молекуле ДНК комплементарны друг другу. Последовательность нуклеотидов в одной из спиралей определяет последовательность нуклеотидов в другой.
В каждой паре оснований, связанных водородными связями, одно из оснований – пуриновое, а другое пиримидиновое. Общее число остатков пуриновых оснований в молекуле ДНК равно числу остатков пиримидиновых оснований.
Таким образом,
- ТИМИН (Т) комплементарен АДЕНИНУ (А),
- ЦИТОЗИН (Ц) комплементарен ГУАНИНУ (Г).
Комплементарность полинуклеотидных цепей служит химической основой главной функции ДНК – хранения и передачи наследственных признаков.
Репликация ДНК
Двухспиральная структура ДНК с комплементарными полинуклеотидными цепями обеспечивает возможность самоудвоения (репликации) этой молекулы.
Перед удвоением водородные связи разрываются, и две цепи раскручиваются и расходятся. Каждая цепь затем служит матрицей для образования на ней комплементарной цепи.
После разделения цепей происходит саморепликация, т.е. образование новой двойной спирали, идентичной исходной.
После репликации образуются две дочерние молекулы ДНК, в каждой из которых одна спираль взята из родительской ДНК, а другая (комплементарная) синтезирована заново.
Таким образом, сохраняется и передается новому поколению исходная структура ДНК.
Длина полинуклеотидных цепей ДНК практически неограничена. Число пар оснований в двойной спирали может меняться от нескольких тысяч у простейших вирусов до сотен миллионов у человека.
Видеофильм «ДНК. Код Жизни»
Рубрики: Нуклеиновые кислоты
Окончательная компактизация ДНК
В ядре форма макромолекулы дезоксирибонуклеиновой кислоты становится чрезвычайно сложной, компактизируясь в несколько этапов.
- Во-первых, нить сворачивается в особую структуру типа соленоида – хроматиновую фибриллу толщиной в 30 нм. На этом уровне ДНК, сворачиваясь, сокращает свою длину в 6-10 раз.
- Далее фибрилла при помощи специфических скэффолд-белков образует зигзагообразные петли, что уменьшает линейный размер ДНК уже в 20-30 раз.
- На следующем уровне формируются плотно упакованные петельные домены, чаще всего имеющие форму, условно названную «ламповая щетка». Они прикрепляются к внутриядерному белковому матриксу. Толщина таких структур составляет уже 700 нм, ДНК при этом укорачивается приблизительно в 200 раз.
- Последний уровень морфологической организации – хромосомный. Петельные домены уплотняются настолько, что достигается общее укорочение в 10 000 раз. Если длина растянутой молекулы – около 5 см, то после упаковки в хромосомы она уменьшается до 5 мкм.
Высшего уровня усложнения формы ДНК достигает в состоянии метафазы митоза. Именно тогда она приобретает характерный облик – две хроматиды, соединенные перетяжкой-центромерой, которая обеспечивает расхождение хроматид в процессе деления. Интерфазная ДНК организована до доменного уровня и распределяется в ядре клетки без особого порядка. Таким образом, мы видим, что морфология ДНК тесно связана с различными фазами ее существования и отражает особенности функционирования этой важнейшей для жизни молекулы.
Как происходит репликация
Матричный синтез ДНК происходит с участием множества ферментов и вспомогательных белков. Ключевыми компонентами являются:
- ДНК-хеликаза — расплетает двойную спираль, разрушает связи между цепями молекулы;
- ДНК-лигаза — «зашивает» разрывы между фрагментами Оказаки;
- праймаза — синтезирует затравку, необходимую для работы ДНК-синтезирующего фрагмента;
- SSB-белки — стабилизируют одноцепочечные фрагменты расплетенной ДНК;
- ДНК-полимеразы — синтезируют дочернюю матричную цепь.
Хеликаза, праймаза и SSB-белки подготавливают почву для синтеза. В результате каждая из цепей исходной молекулы становится матрицей. Синтез осуществляется с огромной скоростью (от 50 нуклеотидов в секунду).
Работа ДНК-полимеразы происходит в направлении от 5`к 3`- концу. Из-за этого на одной из цепей (лидирующей) синтез происходит по ходу расплетания и непрерывно, а на другой (отстающей) — в обратном направлении и отдельными фрагментами, названными «Оказаки».
Y-образная структура, образованная в месте расплетания ДНК, называется репликационной вилкой.
Трансляция
Матричный синтез белка и у прокариот, и у эукариот осуществляется в специализированных органоидах — рибосомах. Последние состоят из двух субъединиц, одна из которых (малая) служит для связывания тРНК и матричной РНК, а другая (большая) принимает участие в образовании пептидных связей.
Началу трансляции предшествует активация аминокислот, т. е. присоединение их к соответствующим транспортным РНК с образованием макроэргической связи, за счет энергии которых впоследствии осуществляются реакции транспептидирования (присоединения к цепи очередного звена).
В процессе синтеза также принимают участие белковые факторы и ГТФ. Энергия последнего необходима для продвижения рибосомы по матричной цепи РНК.
Сущность генетического кода
Итак, ДНК – это информационная матрица, на которой сохраняется информация о необходимых организму для роста и жизнедеятельности белках. Белки строятся из аминокислот, ДНК (и РНК) – из нуклеотидов. Определенным нуклеотидным последовательностям молекулы ДНК соответствуют определенные последовательности аминокислот тех или иных белков.
Структурных единиц белка – канонических аминокислот – в клетке 20 видов, а нуклеотидов в составе ДНК – 4 вида. Так что каждая аминокислота записана на ДНК-матрице как сочетание трех нуклеотидов – триплет, ключевыми компонентами которого являются азотистые основания. Такой принцип соответствия называется генетическим кодом, а триплеты оснований – кодонами. Ген – это последовательность кодонов, содержащая запись какого-либо белка и некоторые служебные сочетания оснований – старт-кодон, стоп-кодон и прочие.
Как воспроизводится генетическая программа
Благодаря комплементарному строению возможно матричное самовоспроизведение молекулы ДНК. При этом происходит копирование содержащейся в ней информации. Удвоение молекулы с образованием двух дочерних «двойных спиралей» называется репликацией ДНК. Это сложный процесс, протекающий с участием многих компонентов. Но при известном упрощении его можно представить в виде схемы.
Инициируется репликация особым комплексом ферментов на определенных участках ДНК. При этом двойная цепочка расплетается, формируя репликационную вилку, где и проходит процесс биосинтеза ДНК – наращивания комплементарных последовательностей нуклеотидов на каждой из цепочек.
Функции ДНК
Рассмотрим, какую роль играет ДНК в живых системах. Этот биополимер является матрицей, содержащей запись структуры различных нужных организму белков, РНК, а также разного рода регуляторных участков. В целом, все эти компоненты составляют генетическую программу организма.
Посредством биосинтеза ДНК генетическая программа передается следующим поколениям, обеспечивая наследственность основополагающей для жизни информации. ДНК способна мутировать, благодаря чему возникает изменчивость живых организмов одного биологического вида и, как следствие, возможен процесс естественного отбора и эволюция живых систем.
При половом размножении ДНК организма-потомка формируется путем сочетания отцовской и материнской наследственной информации. При комбинации ее возникают различные варианты, что также способствует изменчивости.
Транскрипция и трансляция
Это два наиглавнейших шага биосинтеза.
Транскрипция с латинского означает «переписывание» – в качестве матрицы применяется ДНК, поэтому происходит синтезирование трёх видов РНК (матричной/информационной, транспортной, рибосомной рибонуклеиновых кислот). Реакция осуществляется с помощью полимеразы (РНК) и с использованием большого количества аденозинтрифосфата.
Выделают два основных действия:
- Обозначение конца и начала трансляции присоединением иРНК.
- Событие, осуществляемое благодаря сплайсингу, который в свою очередь удаляет неинформационные последовательности РНК, тем самым происходит уменьшение массы матричной рибонуклеиновой кислоты в 10 раз.
Трансляция с латинского означает «перевод» – используется иРНК в качестве матрицы, синтезируются полипептидные цепочки.
Трансляция включает в себя три этапа, которые можно было представить в виде таблицы:
- Первый этап. Инициация формирование комплекса, который участвует в синтезе полипептидной цепочки.
- Второй этап. Элонгация увеличение размеров этой цепи.
- Третий этап. Терминация заключение выше упомянутого процесса.
Обмен веществ
Обмен веществ — важнейшее свойство живых организмов. Совокупность реакций обмена веществ, протекающих в организме, называется метаболизмом. Метаболизм состоит из реакций ассимиляции (пластического обмена, анаболизма) и реакций диссимиляции (энергетического обмена, катаболизма). Ассимиляция — совокупность реакций биосинтеза, протекающих в клетке, диссимиляция — совокупность реакций распада и окисления высокомолекулярных веществ, идущих с выделением энергии. Эти группы реакций взаимосвязаны: реакции биосинтеза невозможны без энергии, которая выделяется в реакциях энергетического обмена, реакции диссимиляции не идут без ферментов, образующихся в реакциях пластического обмена.
По типу обмена веществ организмы подразделяются на две группы: автотрофы и гетеротрофы. Автотрофы — организмы, способные синтезировать органические вещества из неорганических и использующие для этого синтеза или солнечную энергию, или энергию, выделяющуюся при окислении неорганических веществ. Гетеротрофы — организмы, использующие для своей жизнедеятельности органические вещества, синтезированные другими организмами. В качестве источника углерода автотрофы используют неорганические вещества (СО2), а гетеротрофы — экзогенные органические. Источники энергии: у автотрофов — энергия солнечного света (фотоавтотрофы) или энергия, выделяющаяся при окислении неорганических соединений (хемоавтотрофы), у гетеротрофов — энергия окисления органических веществ (хемогетеротрофы).
Большинство живых организмов относится или к фотоавтотрофам (растения), или к хемогетеротрофам (грибы, животные). Если организмы, в зависимости от условий, ведут себя как авто- либо как гетеротрофы, то их называют миксотрофами (эвглена зеленая).
Нуклеиновые кислоты – полимерные молекулы
Нуклеиновые кислоты — самые крупные нерегулярные полимерные органические молекулы, носящие название полинуклеотидов. Обычно ДНК намного крупнее РНК. Их мономерами являются нуклеотиды (нуклеозиды, дезоксинуклеозиды и др.). Каждый из них состоит из трёх компонентов:
- пентозы, или пятиуглеродного сахара (рибоза в РНК и дезоксирибоза в ДНК);
- фосфатной группы – остатка фосфорной кислоты (—PO 4 -);
- азотистого основания.
Строение нуклеотида
Азотистые основания — это ароматические гетероциклические соединения, производные пиримидина или пурина. Нуклеотиды имеют пять основных типов азотистых оснований. Двухкольцевые пуриновые: аденин (Аde) и гуанин (Gua). Каждое из них содержится как в ДНК, так и в РНК. Остальные три основания представляют собой однокольцовые молекулы, производные пиримидина: цитозин (Cyt — есть как в ДНК, так и в РНК), тимин (Thy — только в ДНК), урацил (Ura — только в РНК).
Аденин и рибоза образуют нуклеозид аденозин (A), производные других азотистых оснований носят названия: гуанозин (G, Г), уридин (U, У), тимидин (Т), цитидин (C, Ц). При соединении азотистых оснований с дезоксирибозой образуются дезоксинуклеозиды. Все нуклеозидфосфаты объединяют под общим названием — нуклеотиды.
Строение пурина и пуриновых азотистых основанийСтроение пиримидина и пиримидиновых азотистых оснований
Нуклеиновые кислоты образуются путём реакции обезвоживания (конденсации, или дегидрации) между фосфатной группой одного нуклеотида и гидроксильной группой пентозы другого нуклеотида. Так получается фосфодиэфирная связь, объединяющая два углевода через фосфат.
В молекуле нуклеотида азотистое основание присоединено к первому атому углерода пентозы, а остаток фосфорной кислоты — к пятому. Получающаяся полинуклеотидная цепь полярна, она имеет два конца:
- 5′ (пять-штрих положение) — углеродный атом в пятичленном моносахариде — рибозе или дезоксирибозе;
- 3´ (три-штрих положение) — гидроксильная группа, взятая от углевода (ОН).
Эти концы в двойной спирали ДНК соединяются через фосфатную группу по типу голова-хвост (3′ к 5′) по принципу комплементарности, азотистыми основаниями внутрь спирали. Такая ориентация цепей называется антипараллельной.
Строение и функции РНК
РНК — полимер, мономерами которой являются рибонуклеотиды. В отличие от ДНК, РНК образована не двумя, а одной полинуклеотидной цепочкой (исключение — некоторые РНК-содержащие вирусы имеют двухцепочечную РНК). Нуклеотиды РНК способны образовывать водородные связи между собой. Цепи РНК значительно короче цепей ДНК.
Мономер РНК — нуклеотид (рибонуклеотид) — состоит из остатков трех веществ: 1) азотистого основания, 2) пятиуглеродного моносахарида (пентозы) и 3) фосфорной кислоты. Азотистые основания РНК также относятся к классам пиримидинов и пуринов.
Пиримидиновые основания РНК — урацил, цитозин, пуриновые основания — аденин и гуанин. Моносахарид нуклеотида РНК представлен рибозой.
Выделяют три вида РНК: 1) информационная (матричная) РНК — иРНК (мРНК), 2) транспортная РНК — тРНК, 3) рибосомная РНК — рРНК.
Все виды РНК представляют собой неразветвленные полинуклеотиды, имеют специфическую пространственную конформацию и принимают участие в процессах синтеза белка. Информация о строении всех видов РНК хранится в ДНК. Процесс синтеза РНК на матрице ДНК называется транскрипцией.
Транспортные РНК содержат обычно 76 (от 75 до 95) нуклеотидов; молекулярная масса — 25 000–30 000. На долю тРНК приходится около 10% от общего содержания РНК в клетке. Функции тРНК: 1) транспорт аминокислот к месту синтеза белка, к рибосомам, 2) трансляционный посредник. В клетке встречается около 40 видов тРНК, каждый из них имеет характерную только для него последовательность нуклеотидов. Однако у всех тРНК имеется несколько внутримолекулярных комплементарных участков, из-за которых тРНК приобретают конформацию, напоминающую по форме лист клевера. У любой тРНК есть петля для контакта с рибосомой (1), антикодоновая петля (2), петля для контакта с ферментом (3), акцепторный стебель (4), антикодон (5). Аминокислота присоединяется к 3′-концу акцепторного стебля. Антикодон — три нуклеотида, «опознающие» кодон иРНК. Следует подчеркнуть, что конкретная тРНК может транспортировать строго определенную аминокислоту, соответствующую ее антикодону. Специфичность соединения аминокислоты и тРНК достигается благодаря свойствам фермента аминоацил-тРНК-синтетаза.
Рибосомные РНК содержат 3000–5000 нуклеотидов; молекулярная масса — 1 000 000–1 500 000. На долю рРНК приходится 80–85% от общего содержания РНК в клетке. В комплексе с рибосомными белками рРНК образует рибосомы — органоиды, осуществляющие синтез белка. В эукариотических клетках синтез рРНК происходит в ядрышках. Функции рРНК: 1) необходимый структурный компонент рибосом и, таким образом, обеспечение функционирования рибосом; 2) обеспечение взаимодействия рибосомы и тРНК; 3) первоначальное связывание рибосомы и кодона-инициатора иРНК и определение рамки считывания, 4) формирование активного центра рибосомы.
Информационные РНК разнообразны по содержанию нуклеотидов и молекулярной массе (от 50 000 до 4 000 000). На долю иРНК приходится до 5% от общего содержания РНК в клетке. Функции иРНК: 1) перенос генетической информации от ДНК к рибосомам, 2) матрица для синтеза молекулы белка, 3) определение аминокислотной последовательности первичной структуры белковой молекулы.
А-ДНК – сухая молекула
А-форма – это правый винт с 11 комплементарными парами оснований в каждом витке. Диаметр его составляет 2,3 нм, а длина одного витка спирали – 2,5 нм. Плоскости, образуемые спаренными основаниями, имеют наклон 20° по отношению к оси молекулы. Соседние нуклеотиды расположены в цепочках компактно – между ними всего 0,23 нм.
Такая форма ДНК возникает при низкой гидратации и при повышенной ионной концентрации натрия и калия. Она характерна для процессов, в которых ДНК образует комплекс с РНК, поскольку последняя не способна принимать иные формы. Кроме того, А-форма весьма устойчива к ультрафиолетовому облучению. В этой конфигурации дезоксирибонуклеиновая кислота содержится в грибных спорах.
Механизм транскрипции
Ключевым ферментом транскрипции является РНК-полимераза. Последняя бывает нескольких видов и отличается по строению у прокариот и эукариот. Однако механизм ее действия везде одинаков и заключается в наращивании цепи комплементарно подбираемых рибонуклеотидов с замыканием фосфодиэфирной связи между ними.
Матричной молекулой для этого процесса служит ДНК. На ее основе могут создаваться разные типы РНК, а не только информационные, которые используются в белковом синтезе.
Участок матрицы, с которого «списывается» последовательность РНК, называется транскриптоном. В его составе имеется промотор (место для присоединения РНК-полимеразы) и терминатор, на котором синтез останавливается.
Основные аспекты синтеза молекул на основе матрицы
Информация о структуре синтезируемой молекулы содержится в последовательности звеньев самой матрицы, к каждому из которых подбирается соответствующий элемент «дочерней» цепи. Если химическая природа синтезируемой и матричной молекул совпадают (ДНК-ДНК или ДНК-РНК), то сопряжение происходит напрямую, так как каждый нуклеотид имеет пару, с которой может связаться.
Для синтеза белка требуется посредник, одна часть которого взаимодействует с матрицей по механизму нуклеотидного соответствия, а другая присоединяет белковые звенья. Таким образом, принцип комплементарности нуклеотидов работает и в этом случае, хоть и не связывает напрямую звенья матричной и синтезируемой цепей.
Репликация ДНК и ее строение
И первичная, и вторичная структуры ДНК возникают в ходе явления, называемого репликацией – образования из материнской макромолекулы двух идентичных ей «двойных спиралей». При репликации исходная молекула расплетается, и на освободившихся одиночных цепочках происходит наращивание комплементарных оснований. Поскольку половинки ДНК антипараллельны, этот процесс протекает на них в разных направлениях: по отношению к материнским цепочкам от 3’-конца к 5’-концу, то есть новые цепочки растут в направлении 5’ → 3’. Лидирующая цепь синтезируется непрерывно в сторону репликационной вилки; на отстающей цепи синтез совершается от вилки отдельными участками (фрагменты Оказаки), которые затем сшивает между собой особый фермент – ДНК-лигаза.
Пока продолжается синтез, уже сформированные концы дочерних молекул претерпевают винтообразное закручивание. Затем, еще до окончания репликации новорожденные молекулы начинают образовывать третичную структуру в процессе, именуемом сверхспирализацией.